查吉尔斯卡亚洞穴中一个高覆盖率的尼安德特人基因组
Svante Pääbo,2020年5月13日贡献(2020年3月16日发送供审阅;Kelley Harris和Roberto Macchiarelli审阅)。
我们对来自俄罗斯阿拉泰山脉查盖尔斯卡亚洞穴的一名尼安德特人的基因组进行了测序,基因组覆盖率为27倍。我们表明,这名尼安德塔尔人是一名女性,她与欧亚大陆西部的尼安德塔尔人[Prüfer等人,Science 358,655-658(2017);Hajdinsak等人,Nature 555,652-656(2018)]的亲缘关系比与更早居住在Denisova洞穴的尼安德特人[Prüfer等人,Nature 505,43-49(2014)]的亲缘关系更近。Chagyrskaya约12.9%的基因组由2.5到10厘米长的纯合子区域跨越。这与西伯利亚尼安德特人生活在相对与世隔绝的不到60人的群体中的事实是一致的。相比之下,来自欧洲的尼安德特人,来自阿拉泰山脉的丹尼索瓦人,以及古代现代人似乎生活在更大的群体中。三个高质量的尼安德特人基因组的可获得性使得人们可以看到尼安德特人独有的、可能在他们中出现频率很高的遗传特征。我们发现,在大脑基底节的纹状体中高表达的基因比在大脑其他地方表达的基因携带更多的氨基酸变化替代,这表明纹状体可能在尼安德特人中进化出了独特的功能。
尼安德特人和丹尼索瓦人是现代人在进化上的近亲。对它们基因组的分析表明,它们对撒哈拉以南非洲以外的现代人有遗传贡献(1,2)。然而,到目前为止,只有两个尼安德特人和一个丹尼索瓦人的基因组测序达到了高质量。其中一个尼安德特人基因组(Vindja33.19)来自在克罗地亚Vindja洞穴中发现的一个个体(3),而另一个尼安德特人基因组(Denisova5或“阿拉泰尼安德塔尔人”)(4)和丹尼索瓦人基因组(Denisova3)(5)都来自在西伯利亚阿拉泰山脉的丹尼索瓦洞穴发现的标本。
一些中等质量的古代基因组(一到三倍的基因组复盖率)为尼安德特人的历史提供了更多的洞察力。例如,来自欧洲的5名晚期尼安德特人的基因组序列表明,他们携带的遗传变异很小(6,7),与文迪亚33.19人的亲缘关系比与丹尼索瓦5尼安德特人的亲缘关系更近。来自丹尼索瓦洞穴的一块形态未诊断的骨头(丹尼索瓦11号)的基因组序列属于一位尼安德特人母亲和一位丹尼索瓦人父亲的直系后代(8),表明这两个群体在阿拉泰地区相遇。丹尼索瓦11号的尼安德特人母亲与Vindja 33.19的亲缘关系比与丹尼索瓦5号的亲缘关系更近,表明阿拉泰山脉的尼安德特人种群发生了更替(8)。
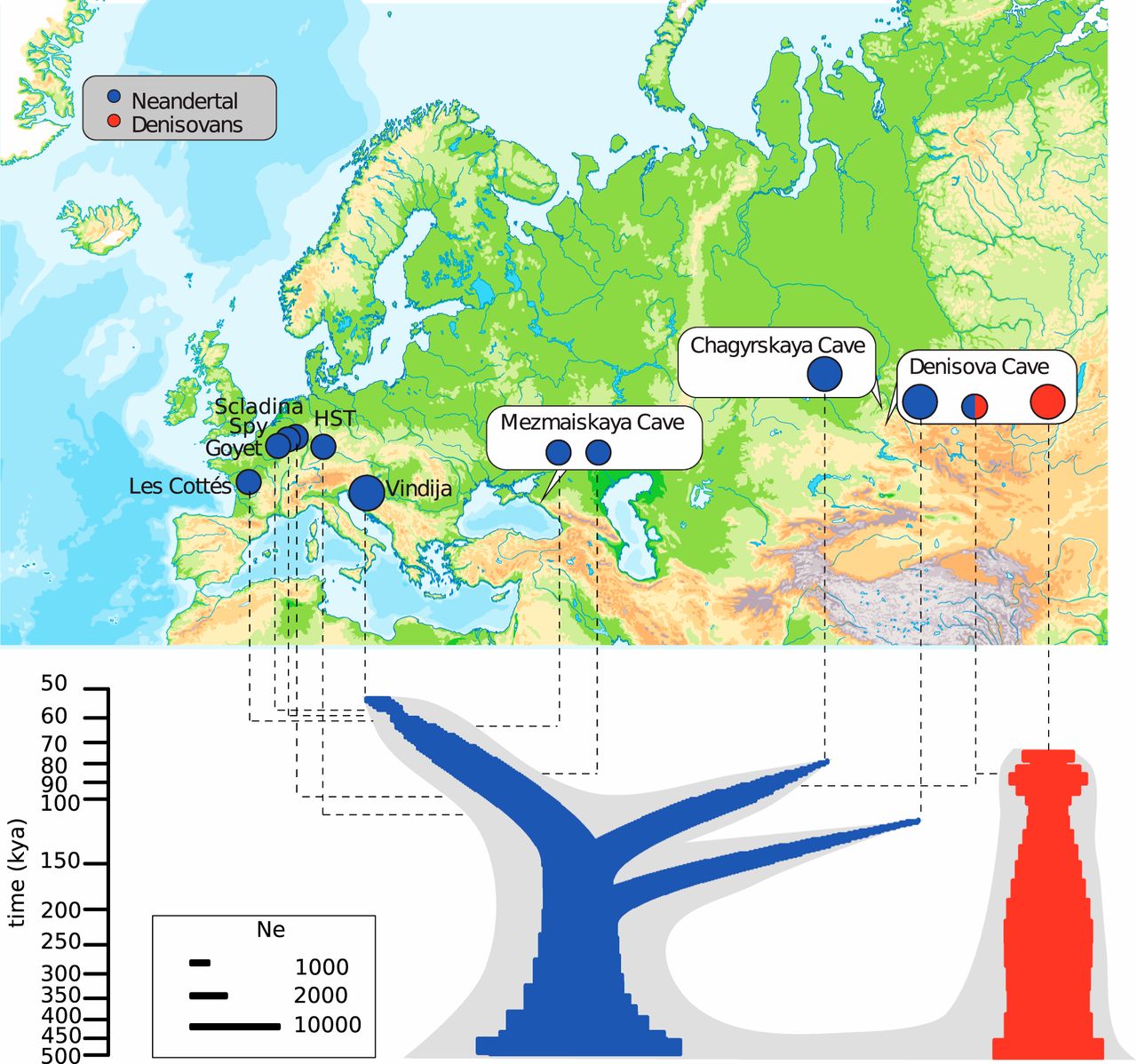
在这里,我们展示了来自查盖尔斯卡亚洞穴的尼安德特人的高覆盖率基因组序列,位于丹尼索瓦洞穴以西106公里处(9⇓⇓-12)(图1)。这个基因组提供了对尼安德特人群体结构和历史的洞察,并允许识别尼安德特人特有的基因组特征。
我们从Chagyrskaya 8采集了100毫克骨粉,这是2011年在6b层的Chagyrskaya洞穴发现的一个方阵(SI附录1)。提取的DNA(14)允许对核基因组进行测序(SI附录2),平均覆盖率为27.6倍(SI附录3)。据估计,测序的DNA片段中只有不到1%来自于现代人类DNA的污染(SI附录4和5)。
我们使用两种不同的方法估计了Chagyrskaya 8的年龄(SI附录6)。首先,我们统计了“缺失的”衍生替代与现在基因组的比例(3个⇓-5,15个)。我们还使用了与Fu等人类似的方法。(15)这利用了三个高覆盖率的尼安德特人基因组的共同进化史。在尼安德塔尔人与现代人具有相同的突变率(每代每碱基对1.45×10−8突变)(15)和世代时间(29y)的情况下,两种方法都表明Chagyrskaya 8生活在∼80kya(1000年前),即Denisova5之后的∼30Ky(1000y)和∼30Ky之前的Vindja33.19。这一估计比发现查吉尔斯卡亚8的考古层的∼60kya(10,12)的光刺激发光日期要早。排除来自较低的、较老的层(10,12)的再沉积,这可能表明基于目前人类突变率的遗传日期是不正确的。可能的解释是,尼安德特人的突变率比现代人低,或者现代人的突变率最近有所下降(16)。为了解决这些可能性,需要从年代明确的尼安德特人遗骸中确定更多的高质量基因组。然而,Chagyrskaya 8尼安德特人和Denisovan Denisova 3显示出相似的比例
与Vindja33.19(图2)相比,Chagyrskaya 8与生活在欧洲∼50kya的其他尼安德特人分享的派生等位基因较少,即与Vindja 33.19大致相同。然而,Chagyrskaya 8与第一代尼安德塔尔-丹尼索瓦人后代Denisova 11的衍生等位基因比Vindja 33.19共享更多的等位基因(8)。由于Vindja 33.19和Chagyrskaya 8与Denisovan Denisova 3在派生等位基因上没有不同,这表明Chagyrskaya 8在目前已知的Neandertal中与在Denisova洞穴中发现的Denisova 11的母亲关系最密切(图1和SI附录7)。
非非洲现代人携带∼2%(1,3)尼安德特人祖先,这是来自尼安德特人的基因流动的结果,发生在50到90 kya(1,17)之间。在全基因组范围内,Chagyrskaya 8与非洲以外的现代人类群体的等位基因比Denisova 5多,等位基因比例与Vindja 33.19相似(SI附录7)。然而,如果分析局限于之前检测到的导入到现代人中的尼安德特人单倍型(18),或者局限于在今天的非非洲人群中出现频率较低的、因此更有可能从尼安德特人导入的派生等位基因,那么Vindja 33.19与今天的人群分享的等位基因比Chagyrskaya 8更多。这表明Vindja 33.19比Chagyrskaya 8与贡献过的尼安德特人群体的亲缘关系更密切。这就是说,Vindja 33.19比Chagyrskaya 8更接近于对尼安德特人群体有贡献的尼安德特人群体,Vindja 33.19与Chagyrskaya 8的关系比Chagyrskaya 8更密切。
为了测试是否有任何现代人群体携带来自尼安德特人的额外基因贡献,这些尼安德特人与查吉尔斯卡娅8号的亲缘关系比与Vindja 33.19的亲缘关系更近,我们利用了之前发表的单倍型推断,这些单倍型来自尼安德特人,这些单倍型今天只在东亚、欧洲、印度或大洋洲发现。在来自西蒙斯基因组多样性小组(19)的300个基因组和89个巴布亚基因组(20,21)中,与Chagyrskaya 8和Vindja 33.19共享的等位基因比例相似(SI附录7),没有迹象表明尼安德特人与Chagyrskaya 8的亲缘关系比与Vindja 33.19的亲缘关系更近。因此,在这些分析的解决范围内,我们得出结论,如果几个尼安德特人群体以不同的比例(22,23)对不同现代人群体的基因组做出贡献,这些尼安德特人群体与Chagyrskaya 8和Vindja 33.19相似。
丹尼索瓦洞穴的尼安德特人基因组,丹尼索瓦5号,携带着很高比例的遗传纯合的长链(HBD链)(4)。超过10厘米的管束表明丹尼索瓦5号的父母是近亲,而2.5厘米至10厘米的HBD管束表明丹尼索瓦5号来自的种群规模较小,在个体生活之前的100代人中,丹尼索瓦5号的种群规模较小(SI附录8)。
与Denisova 5相比,Chagyrskaya 8基因组携带的10 cm以上的HBD区较少,但中等长度的HBD区较多(SI附录8)。事实上,所有三个高覆盖率的尼安德特人基因组都携带了比几乎所有现代和史前现代人类基因组以及丹尼索瓦人基因组(Denisova 3)更多的中等大小的HBD区域(图3A)。我们通过联合模拟表明,这不能用总体规模较小但泛滥的人口来解释。相反,它表明尼安德特人的种群是细分的(SI附录8)。
通过联合建模,我们推测Chagyrskaya 8和Denisova 5可能生活在60个或更少的亚群中。相反,现在和过去的现代人群体以及丹尼索瓦人(基于Denisova3基因组)生活在100多个个体的子群体中,假设子群体之间的迁移率为1%或更少(图3B和SI附录8)。有趣的是,Vindja 33.19尼安德特人似乎生活在一个比两个西伯利亚尼安德特人更大的亚群中,尽管当考虑到所有HBD束覆盖的基因组比例超过2.5厘米时,这种差异在统计上只有轻微的显著性(似然比检验,P=0.05)。
我们使用三个高复盖率的尼安德特人基因组来识别生物学途径,其中蛋白质编码基因在三个尼安德特人中表现出比预期更多的非同义替换,这是因为沉默和多态的变化。我们在889个基因中确定了993个固定的非同义替换,在3个尼安德特人中发现了2952个多态的非同义替换。
与其他基因组(家族错误率0.1)相比,没有与特定已知生物学功能或表型相关的基因组(26)相对于非同义与同义多态改变的比率(麦克唐纳-克莱特曼比率)(27)显示出较高的非同义固定改变与同义固定改变的比率(28和SI附录9)。
然而,当根据Allen脑图谱(29,30)分析在不同脑区优先表达的基因时,我们发现在12-19岁的个体中,纹状体表达的基因的麦克唐纳-克莱特曼比(1.0 2,家族错误率=0.029)高于在其他脑区和其他年龄表达的基因(0.5 3-0.83)(SI附录9),但在12-19岁的个体中,纹状体表达的基因显示出更高的麦克唐纳-克莱特曼比(1.0 2,家族错误率=0.5 3)(SI附录9)。这可能表明,在尼安德特人中,纹状体表达的基因的负选择被放松了。或者,这些基因编码的一些蛋白质可能是正选择的目标。此外,在胎儿期纹状体表达的基因在其非翻译区比在其他地方表达的基因携带更多的替换(家族错误率=0.049)和其他时间。在纹状体表达的基因中,那些携带尼安德特人固定的非同义变化的基因比不携带这种变化的纹状体基因更常出现在现代人类中携带很少或没有从尼安德特人导入的dna的基因组区域(费舍尔精确检验P=0.026)。在尼安德塔尔人(P&gT;0.1)中,并不是所有携带固定非同义替换的基因都观察到了这种模式,这表明在现代人中,尼安德特人纹状体基因中的一些替换可能是负选择的。除了纹状体外,在尼安德特人中,产前在顶后皮质、腹外侧额前皮质和初级躯体感觉皮层表达的基因在它们的调节区比在其他脑区和其他时间表达的基因具有更多的固定替换(SI附录9)。在尼安德特人中,除了纹状体外,在顶叶后皮质、腹外侧前额叶皮质和初级躯体感觉皮层表达的基因在它们的调节区具有更多的固定替换(SI附录9)。
为了进一步检测沿着尼安德特人血统的阳性选择,我们在基因组的25kb滑动窗口中进行了Hudson,Kreitman,Aguadé(HKA)测试(31)和种群分支统计(PBS)(32)(SI附录10)。我们估计了通过合并模拟获得不同统计数据的观测值的可能性,并保留了错误发现率为5%的窗口(图4)。在常染色体上,我们总共识别了35个不同的候选区。其中一个候选区域,由第5号染色体上的PBS确定的75kb长的区域,与HKA确定的两个单独的窗口重叠。此重叠低于更改后的预期(P值<;4×10−4和SI附录10)。候选区域确定了与神经发育(EXOC6B)、免疫、伤口愈合(HTN1、EVPLL)和线粒体功能(NSUN3、TIMM29)相关的重叠基因。对于这两个统计数据,我们发现与以前在现代人中被确认为正选择的基因组区域(33)有重叠(富集测试,香港A和PBS的P值分别为0.010和0.056)。
Chagyrskaya 8与文迪亚33.19和其他欧亚大陆西部的其他晚期尼安德特人的亲缘关系比与较早生活在阿拉泰山脉的丹尼索瓦5尼安德特人的亲缘关系更近(图2)。因此,Chagyrskaya 8与在120到80 kya之间向东迁移的尼安德特人有关(13)。有趣的是,Chagyrskaya洞穴中发现的文物与中欧和东欧的文物组合有相似之处(10和SI附录1),这表明从西欧来到西伯利亚的尼安德特人可能带来了他们的物质文化(10,34)。这些到来的尼安德特人中有一些遇到了当地的丹尼索瓦人,正如丹尼索瓦11岁所展示的那样,他的父亲是丹尼索瓦人,母亲是尼安德特人,与Chagyrskaya 8人生活的人口有血缘关系。
在这方面,有趣的是,Chagyrskaya 8和Denisova 5尼安德塔尔人的种群比克罗地亚的Vindja 33.19尼安德塔尔人、Denisovan Denisova 3人和现代人(24人)要少(图3B)。阿拉泰地区的尼安德特人可能比其他地方的尼安德特人生活在更小、更与世隔绝的人口中,因为该地区代表了他们地理分布的外围,可能是丹尼索瓦人更持续存在的地区。然而,需要对丹尼索瓦洞穴和其他遗址的人口历史进行更详细的研究才能澄清这一点。
当使用三个尼安德特人基因组分析尼安德特人血统的遗传变化时,青春期纹状体中表达的基因变化的数量尤为突出。一种可能性是,这些变化在尼安德特人身上积累起来,可能是因为他们的种群规模较小,可能与放松了对纹状体表达基因的选择有关。有趣的是,在纹状体表达的基因比预期的更频繁地与基因组区域重叠,而在现代人类基因组中,尼安德特人导入的片段很少。我们推测,纹状体基因可能携带尼安德特人特有的变化,这些变化在引入现代人时是不利的。这一点,以及对现代人血统派生变化的积极选择,可能是目前所谓的尼安德特沙漠的基础。
这项研究由马克斯·普朗克学会、欧洲研究理事会(授予S.P.的赠款协议694707)和俄罗斯科学基金会(项目19-48-04107至K.A.K.)资助。我们感谢Sarah Nagel、Birgit Nickel、Barbara Schellbach和Antje Weihmann的实验室工作,以及Heiko Temming的计算机断层扫描。
作者贡献:K.P.和S.P.设计的研究;F.M.、S.G.、C.D.F.、V.S.、K.A.K.、B.V.、S.V.M.、M.C.、S.P.、L.S.、A.I.K.、A.P.D.、M.M.、J.K.、B.P.、K.P.和S.P.进行研究;L.S.提供新的试剂/分析工具;F.M.、K.A.K.、B.V.、S.V.M.、P.S.、A.I.K.、A.P.D.、M.M.、J.K.、B.P.和K.P.分析了数据;F.M.、A.I.K.、A.P.D.、J.K.、K.P.和S.P.撰写了论文。
资料整理:Chagyrskaya 8的基因组序列可从http://ftp.eva.mpg.de/neandertal/Chagyrskaya/VCF/.下载。其线粒体DNA基因组序列已登录GenBank(登录号:MK388903)。
视图摘要