对100多万人的研究发现寿命与血铁水平有关
衰老表型,如健康状况良好的年数(健康跨度)、总寿命(寿命),以及活到特别高龄(长寿),都是我们都感兴趣的,但需要非常大的样本量来进行遗传学研究。在这里,我们将现有的全基因组范围内关于健康跨度、双亲寿命和寿命的汇总统计数据结合在一个多变量框架中,增加统计能力,并确定影响所有三种表型的10个基因组位点,其中5个(接近FOXO3、SLC4A7、LINC02513、ZW10和FGD6)以前在全基因组意义上尚未报道。这10个基因座中的大多数与心血管疾病有关,有些会影响已知的基因表达,这些基因的活性会随着年龄的增长而改变。我们总共涉及78个基因,并发现这些基因在先前在模式生物中强调的衰老途径中得到了丰富,例如对DNA损伤、凋亡和动态平衡的反应。最后,我们确定了一个值得进一步研究的途径:血红素代谢。
人类衰老的特点是维持体内平衡的能力逐渐下降,导致与年龄相关的疾病的发病,并最终导致死亡。然而,个体之间有很大的差异,有些人很早就患上了慢性病,在60岁之前就去世了,而另一些人则能活到不同寻常的高龄,通常直到生命的最后几年都没有疾病1。健康长寿是由许多不同的因素决定的,包括生活方式、环境、基因和纯粹的机会。最近的估计表明,人类寿命(即活着的年数)和健康跨度(健康无病生活的年限)的基因成分只有10%2,3左右,这使得对这些特征的基因研究具有挑战性,因为除非样本量很大,否则噪音往往会掩盖影响。
然而,有了足够大的样本,对衰老特征的全基因组关联研究(GWAS)就有可能识别参与人类衰老过程的基因和途径。GWAS试图确定与健康寿命3、4、(双亲)寿命5、6、7和超高龄(通常称为长寿)8、9相关的位点和途径,结果之间有一些重叠。相关性状的多变量分析提供了增加力量的前景,特别是在样本不重叠的情况下,他们可能能够识别影响共同潜在衰老过程的变量。
在这里,我们评估了已发表的三种不同类型衰老表型的GWAS之间的基因重叠程度-健康跨度、双亲寿命和寿命(定义为活到90%以上的年龄)-并进行多变量荟萃分析,以确定与健康老龄化相关的遗传变异。随后,我们描述了影响所有这三个衰老性状的基因座的性别和年龄特异性效应,并查找了与年龄相关的表型和疾病的相关报道。最后,我们将这些位点上观察到的信号与特定基因的表达联系起来,包括目前在模式生物中研究的一些基因,并确定与健康衰老有关的途径。
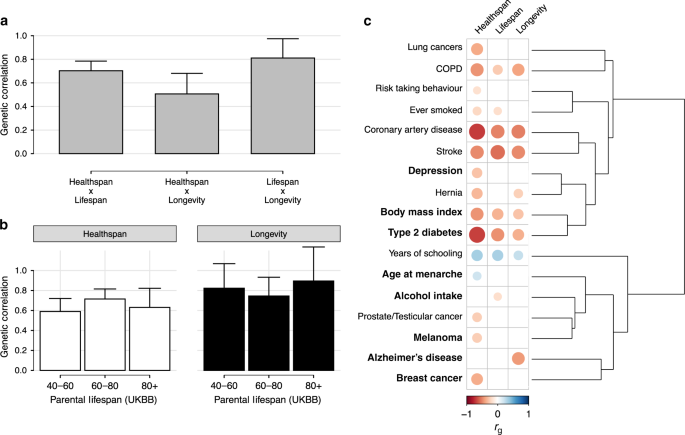
我们探索了三个公共的、欧洲血统的GWAS的重叠衰老特征:健康跨度(N = 300,477人,28.3%不再健康),父母寿命(N = 1,012,240个父母,60%死亡)和长寿(N例 = 11,262;N对照 = 25,483)。尽管在年龄人口统计、性状定义和研究设计上存在差异,但这些性状表现出显著的遗传相关性(P <; 5 × 10-8)。亲本寿命与健康寿命(r g = 0.70;SE = 0.0 4)和寿命(r g = 0.81;SE = 0.0 8)有很强的相关性,而健康寿命和寿命之间的相关性较弱(r g = 0.5 1;SE = 0.0 9)(图1a)。我们在UK Biobank中对双亲寿命进行了年龄分层的GWAS,以评估这些性状之间的遗传相关性是否与年龄有关,但我们的结果显示,健康跨度/寿命与年龄分层的双亲寿命区间之间的相关性没有明显的趋势(图1b)。
人类衰老研究之间的成对遗传相关性。年龄分层双亲寿命与健康寿命和寿命的遗传相关性。C衰老性状与发育、行为和疾病相关性状的遗传相关性(Rg)。粗体是具有异质相关性的性状(Phet <; 005)。在测试的27个性状中,这里显示了17个性状,它们至少有一个显著的遗传相关性(FDR <; 5%)与健康寿命、双亲寿命或长寿有关。根据17个性状与所有被测性状(共30个)的遗传相关性,利用欧几里德距离对其进行聚类。有关相关性的完整列表,请参阅补充数据1;用于计算每个成对相关的SNP数量,请参阅补充表1。空白正方形表示未通过多次的相关性
曼哈顿曲线图显示−在y轴上的标称强度log10(P值)(双面)与SNP在x轴上的染色体位置相对应,其中无效假设与健康寿命、双亲寿命和长寿无关。红线代表全基因组意义阈值(5 × 10-8)。注解的是距离每个位点的前导SNP最近的基因(红色)。为便于辨认,y轴的上限为5 × 10−30;通过此上限的SNP表示为三角形:Lpa,P = 3.8 × 10−30,ApoE,P = 9.6 × 10−127。
在达到全基因组意义的24个多变量GWAS基因座中,有21个在三个研究数据集中具有方向性一致的效应,18个在两个或更多数据集中具有名义上的显著性(P <; 0.05)(补充图1)。在GWAS目录和PhenoScanner中查找铅SNP和接近的替代基因表明,健康跨度特定的基因座(即仅在健康跨度数据集中的P <; 0.05)主要与皮肤癌和代谢特征有关,而父母的寿命特定的基因座与吸烟和冒险行为有关(补充数据3)。与这些表型的关联表明,这些变异会影响(导致)环境暴露的行为,因此很可能捕捉到外在的衰老过程。由于我们主要对影响内在衰老过程的遗传变异感兴趣,所以我们将这项研究的其余部分集中在所有三个数据集中达到名义意义的遗传变异上,这些变异不太可能与研究或特定人群的暴露相关。
在所有三个GWAS数据集中,有10个位点达到名义显著性(P <; 0.0 5)(表1)。其中五个特别令人感兴趣,因为它们在任何单个数据集中都不包含全基因组显着的SNP。这些位点的前导多变量SNP包括rs2643826(最近基因SLC4A7)、rs17499404(LINC02513)、rs1159806(FOXO3)、rs61905747(ZW10)和rs12830425(FGD6)(补充图2-6)。FOXO_3附近的领先单核苷酸多态性与长寿研究13中已知的候选单核苷酸多态性rs2802292处于中度连锁不平衡(r2 >; .4)。考虑到一些位点的P值接近全基因组显著性阈值(即SLC4A7和LINC02513),有必要在我们尚未获得的大的独立队列中复制这些位点。
接下来,我们使用Timmers等人的性别特定的父母GWAS汇总统计数据,测试了感兴趣的位点是否按性别对父母的寿命显示出不同的影响。7.我们发现载脂蛋白Eε4等位基因(βParents = 0.08,βMothers = 0.13,P Diff = 1.5 × 10-6)存在性别二态现象,而LINC02513、SLC4A7、LPA、TOX3和FOXO_3附近的铅SNP没有性别二态现象(<;20%的差异或P Diff >; 为0.50)。对于其余的基因座,在CDKN2B-AS1、ZW10、FGD6和LDLR附近,效应大小点估计可能会有超过20%的差异,但我们需要更大的样本量才能有把握地检测到这种差异(补充图7中)。
在我们的年龄分层的双亲寿命GWAS中查找相同的SNPs,我们发现,除了APOE和SLC4A7外,所有的基因座的有效大小都随着双亲的年龄而呈现下降的趋势。这一趋势对载脂蛋白E位点有显著意义(P调整后的 = 0.0 1),亲本存活率每增加10年,ε4等位基因的效应大小增加32%(2 5-39%)。虽然我们无法确认其余基因座的趋势,但我们发现,总体来说,这9个基因座的保护等位基因的平均效应每增加10年亲本存活率就会降低24%(13-34%;P调整后的 = 1 × 10-4)(补充图8)。
我们还发现,这些感兴趣的基因座以前曾在全基因组显着水平上与几种年龄相关的疾病和/或表型相关。大多数基因座的延长寿命等位基因与心血管疾病表型的减少有关,包括本研究中发现的衰老基因座SLC4A7、FGD6和LINC02513附近的SNP。有趣的是,FOXO3附近的保护性变异与代谢综合征的减少有关,但也与认知能力的降低有关。APOE、FOXO3和FGD6附近的延长寿命的SNP都与黄斑变性的增加有关(补充图9;补充数据3)。
用基因表达数量性状位点(EQTL)评估共定位感兴趣的位点,我们发现10个位点中有8个是顺式作用eQTL共定位的有力证据(FDR SMR <; 5%;P Heidi >; 1%;见“方法”)。总而言之,我们强调了27个独特的基因,作用于32个组织,特别是全血(12个基因)和胫神经(7个基因)(补充数据5)。在血液中,BCL3和CKM(接近APOE)、CTC-510F12.2、ILF3、KANK2和PDE4A(靠近LDLR)、USP28和ANKK1(接近ZW10)和CDKN2B的高表达水平与In
最后,对50条标志性基因和7,350条生物过程途径中的顺式和反式作用基因进行测试,我们发现在7个标志性基因集合和32个生物学过程中显著富集(P调整后的 <; 0.05)。富集程度最高的标志性基因组包括血红素代谢、缺氧和早期雌激素反应(图3)。丰富的生物途径可分为涉及凋亡信号、化学动态平衡、红细胞和髓样细胞发育等类别(补充图10;补充数据7)。
N感兴趣的基因数量与eQTL可用的基因集合中的基因总数之比。浓缩超几何检验的标称P值(对照24,670个背景基因)。P bonf Bonferroni-校正P值,用于测试七条标志性通路(至少包含三个基因)。该图显示了x轴上的单个基因和y轴上列出的标记路径,与表中的顺序一致。方块表示基因集中存在一个基因。
我们假设血红素代谢和化学动态平衡对健康寿命、双亲寿命和寿命的影响可能是通过铁的生物可利用性来介导的,并使用GWAS的MR对铁相关性状的汇总统计数据(即血清铁、对数铁蛋白和转铁蛋白(饱和百分比和绝对水平))与我们的GWAS结果进行了对比,对这一假设进行了研究。在单变量MR框架中,我们发现血清铁(FDR <; 5%)存在因果效应的证据,这似乎与MR的假设一致,对异常值具有鲁棒性(补充图11;补充表2和3)。我们还发现了转铁蛋白饱和效应的一些证据。然而,这种关联主要是由众所周知的遗传性血色素沉着症位点驱动的,并显示出违反多效性假设(即非零MR-Egger截距)的证据。因此,我们在多变量MR分析中测试了所有铁特征作为暴露的同时,以我们的GWAS为结果,发现了血清铁、转铁蛋白水平和转铁蛋白饱和度的因果效应(FDR <; 5%;βIntercept = 0.0012;95%CI:−0.0005至0.0029)的更可靠证据。这些效应并不只由一个基因位点驱动,包括遗传性血色素沉着症基因座,正如留一法分析所证实的那样(补充数据8)。尽管因果效应的单位在不同的暴露中是一致的(并且与P值相关),但它们很难解释。因此,我们对个体成分性状:健康跨度、双亲寿命和寿命重复了这一过程,认识到有效样本量的减少可能会产生动力不足的效应大小估计,尽管这些估计给出了以可测量单位表示的影响的方向和大小(表3)。多变量MR效应的大小似乎比单变量MR的大小更大,这可能是由于动态平衡,即,一个暴露中的变化通常被另一个暴露中的变化缓冲。例如,当金属与转铁蛋白结合时,血清铁的氧化损伤可以在很大程度上得到防止。
公众可获得的健康寿命、双亲寿命和长寿之间的遗传相关性揭示了这些特征共享其潜在遗传学的50%或更多。对GWAS汇总统计数据进行多变量荟萃分析,我们突出了影响一个或多个性状的24个基因组区域。有十个区域特别令人感兴趣,因为它们与所有三个衰老特征相关联,因此有可能捕捉到内在的衰老过程,而不是外部的衰老来源。在任何个体数据集中,包括FOXO3附近的区域,其中五个感兴趣的位点在全基因组显著水平上都没有关联,到目前为止,该区域只在候选基因关联研究中得到了强调(在萨尼斯等人中进行了综述)。15)而且从未在全基因组范围内具有重要意义。兴趣位点对男性和女性寿命的影响大致相同,尽管与老年(80岁)相比,中年(40-60岁)对生存的影响可能略强一些。载脂蛋白Eε4等位基因在这方面是例外的,因为它在女性中的作用更强,并且随着年龄的增长而增加,这可能是因为它与阿尔茨海默病16有众所周知的联系。我们发现这些感兴趣的基因位点与28个顺式基因和50个反式基因的表达是共同定位的,其中一些已知会随着年龄的增长而出现差异表达。最后,我们发现这些基因富集了7组标志性基因(特别是血红素代谢)和32条很大程度上重叠的生物途径(包括凋亡和稳态),并且与突出显示的途径一致,我们在MR框架中发现了健康生活中铁水平的因果作用。
对磁共振结果的解释应谨慎处理,应用方法的透明度以及敏感性分析。