<生>硅藻
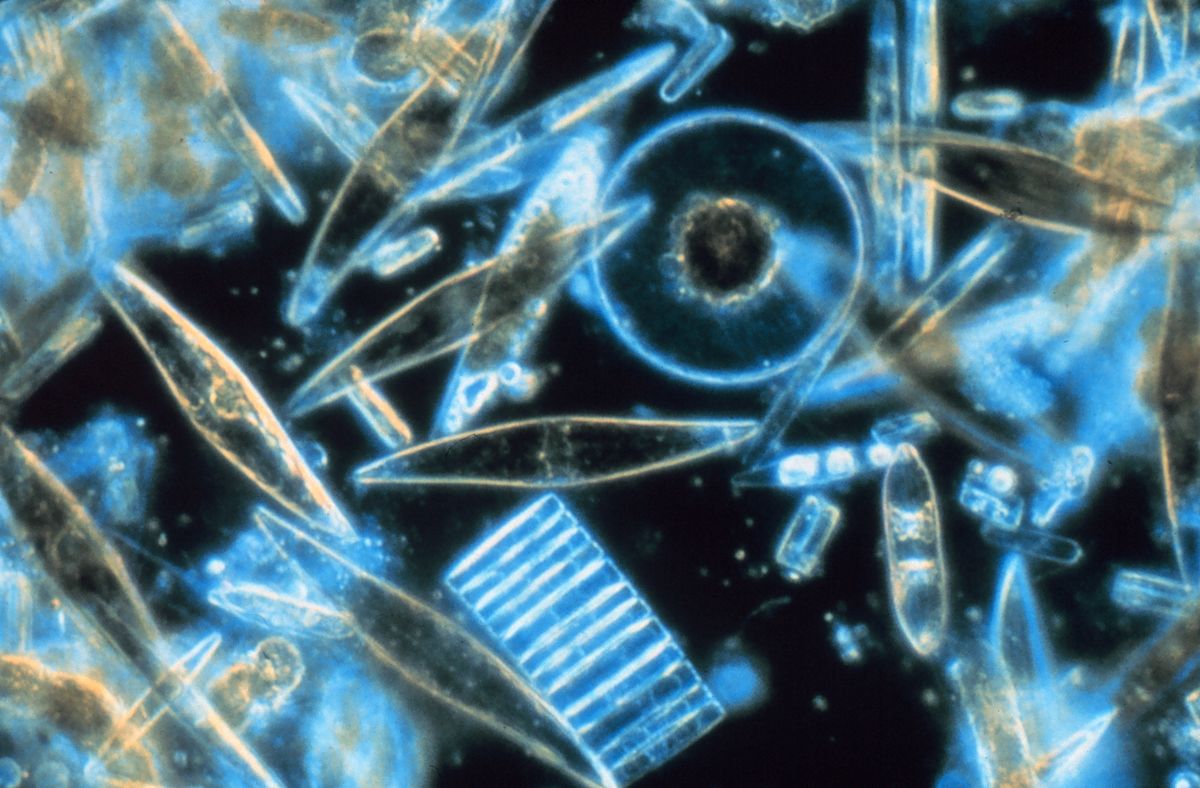
跳转到导航跳转搜索硅藻(Diátom-os';从diá切成两半)[10]是一大类藻类,特别是微藻,发现于世界各地的海洋、水道和土壤中。tém-n-ō的根是一种主要的藻类,[11]特别是微藻,它们存在于世界各地的海洋、水道和土壤中,它们的根部是tém-n-os,i Cut_(39;i)_。活着的硅藻构成了地球生物量的很大一部分:它们每年产生约20%到50%的地球上产生的氧气,[12][13]每年从它们生活的水域中吸收超过67亿公吨的硅,[14]并构成海洋中发现的有机物质的近一半。死亡硅藻的贝壳可以到达海底多达半英里(800米)深的地方,整个亚马逊盆地每年都会受到2700万吨硅藻贝壳灰尘的肥沃,这些灰尘是由来自非洲撒哈拉的大西洋风吹来的,其中大部分来自博代莱凹陷(BodéléDepression),那里曾经是由一个淡水湖泊系统组成的。[15][16]。
硅藻是单细胞的:它们要么以单个细胞的形式出现,要么以群体的形式出现,这些群体的形状可以是丝带、扇形、之字形或星形。单个细胞的大小从2微米到200微米不等。[17]在充足的营养和阳光存在的情况下,通过无性多重分裂,活的硅藻群落大约每24小时翻一番;单个细胞的最大寿命约为6天。[18]硅藻有两种截然不同的形状:少数(中心硅藻)径向对称,而大多数(羽状硅藻)广泛两侧对称。硅藻解剖学的一个独特特征是,它们被由二氧化硅(水合二氧化硅)组成的细胞壁所包围,这种细胞壁被称为锥体。[19]由于它们的光子纳米结构,这些锥体具有结构颜色,这促使它们被描述为海洋的明珠和活的蛋白石。硅藻的运动主要是由于水流和风致水湍流的共同作用而被动发生的,而中心硅藻的雄配子有鞭毛,可以主动运动来寻找雌配子。与植物相似,硅藻通过光合作用将光能转化为化学能,尽管这种共同的自养在两个谱系中都是独立进化的。不同寻常的是,对于自养生物来说,硅藻具有尿素循环,这是它们与动物共有的一个特征,尽管这个循环在硅藻中用于不同的新陈代谢目的。禾本科还拥有一种蓝藻内共生体,称为椭球体。这种内生共生体失去了它的光合作用特性,但保持了它的固氮能力,使硅藻能够固定大气中的氮。[20]。
硅藻研究是生理学的一个分支。硅藻被归类为真核生物,即有膜结合的细胞核的生物体,这将它们与原核生物、古菌和细菌区分开来。硅藻是一种叫做浮游植物的浮游生物,是浮游生物中最常见的一种。硅藻也附着在底栖底物、漂浮的碎屑和大型植物上生长。它们构成了附生植物群落的一个组成部分。[21]另一种分类根据大小将浮游生物分为八类:在这个方案中,硅藻被归类为微藻。存在几种对单个硅藻物种进行分类的系统。化石证据表明,硅藻起源于早侏罗世时期或之前,大约在1.5亿至2亿年前。
硅藻用于监测过去和现在的环境状况,通常用于水质研究。硅藻土(硅藻土)是在地壳中发现的硅藻壳的集合。它们是软的、含二氧化硅的沉积岩,很容易粉碎成细粉,通常粒度为10到200μm。硅藻土可用于各种目的,包括水过滤、作为温和的研磨剂、在猫砂中以及用作炸药稳定剂。
硅藻的长度为2到200微米。[17]它们的黄褐色叶绿体是光合作用的场所,是典型的异形体,有四层膜,含有类胡萝卜素岩藻黄质等色素。个体通常没有鞭毛,但它们存在于中心硅藻的雄配子中,并具有通常的异形体结构,除非它们缺乏其他类群特有的毛发()。
由于其光学特性,硅藻通常被称为海洋之珠或活蛋白石。[22]这种结构着色的生物学功能尚不清楚,但推测可能与通讯、伪装、热交换和/或紫外线防护有关。[23]。
硅藻建造错综复杂、坚硬但多孔的细胞壁,称为主要由硅石组成的锥体。[24]:25-30该硅质壁[25]可以高度图案化,具有各种孔、肋、微孔
大多数中心型和蚜型羽状硅藻是不活动的,它们相对致密的细胞壁使它们很容易下沉。开放水域中的浮游生物形态通常依赖于风对海洋上层的湍流混合,使它们悬浮在阳光充足的水面上。许多浮游硅藻也进化出减缓其下沉速度的特征,如刺或在殖民地链中生长的能力。[26]这些适应增加了它们的表面积与体积比和阻力,使它们能够在水柱中停留更长时间。单个细胞可以通过离子泵调节浮力。[27][27]。
一些羽状硅藻能够进行一种称为滑动的运动,这种运动允许它们通过中缝(阀门表面的一条细长缝隙)分泌的粘液在表面上移动。[28][29]为了使硅藻细胞滑动,它必须有粘液附着的固体底物。
细胞孤立或结合成各种菌落,这些菌落可以通过硅质结构、粘液垫、粘液柄或粘液管、无定形的粘液团或通过细胞突起分泌的甲壳素(多糖)而联系在一起,这些菌落可能是由硅质结构、粘液垫、粘液柄或粘液管、无定形粘液团或几丁质丝(多糖)连接在一起的。
硅藻主要是光合作用的,但也有少数是专性异养生物,只要有合适的有机碳源,它们就可以在没有光的情况下生存。[30][31][31]。
硅藻细胞包含在一种独特的硅质细胞壁中,这种细胞壁被称为锥体,由两个通常相互重叠的瓣膜组成。[32]通过硅酸单体的聚合在细胞内合成构成细胞壁的生物二氧化硅。然后将该材质挤出到单元格外部并添加到墙中。在大多数物种中,当硅藻分裂产生两个子细胞时,每个细胞保留其中一半,并在其内生长较小的一半。因此,在每个分裂周期之后,种群中的硅藻细胞的平均尺寸变小。一旦这些细胞达到一定的最小尺寸,而不是简单地分裂,它们就会通过形成辅助孢子来扭转这种下降趋势。这会在大小上扩大,形成一个大得多的细胞,然后细胞又回到了大小不断缩小的分裂状态。[需要引用]辅助孢子的产生几乎总是与减数分裂和有性繁殖有关。
硅藻吸收的二氧化硅转移到细胞壁的确切机制尚不清楚。硅藻基因的大部分测序来自于研究硅藻在锥体中以纳米尺度模式吸收和沉积的机制。在这一领域取得最大成功的是两个物种,一种是随着全基因组测序和遗传控制方法的建立而成为模式物种的假海链藻,另一种是首次发现重要的硅沉积蛋白silaffins的梭形柱藻(Cylindrotheca Fusiformis)。[33]在梭形芽孢杆菌细胞壁中发现了硅烷烃,这是一组聚阳离子多肽,可以产生复杂的二氧化硅结构。这些结构显示出硅藻图案特有的大小的孔隙。当对假毛虫进行基因组分析时,发现它编码一个尿素循环,包括比大多数基因组更多的多胺,以及三个不同的硅质运输基因。[34]在对8个硅藻不同类群的二氧化硅运输基因的系统发育研究中,发现二氧化硅运输一般按物种归类。[33]这项研究还发现了羽状硅藻(双边对称)和中心硅藻(径向对称)的二氧化硅转运体之间的结构差异。本研究中比较的序列被用来创建不同的背景,以便识别在二氧化硅沉积过程中区分功能的残基。此外,同一项研究发现,许多区域在物种内是保守的,很可能是二氧化硅运输的基础结构。
这些二氧化硅运输蛋白是硅藻独有的,在其他物种中没有发现同源蛋白,如海绵或水稻。这些硅胶转运基因的差异也表明蛋白质的结构是从由五个膜结合片段组成的两个重复单元进化而来的,这表明基因复制或二聚化。[33]硅藻中从膜结合的囊泡中发生的二氧化硅沉积被假设为是由于硅藻中的硅烷和长链多胺的活性所致。这种硅沉积小泡(SDV)的特征是酸性隔室与高尔基体小泡融合在一起。[35]这两种蛋白质结构已被证明在体内产生硅藻小锥尺度上具有不规则孔的图案化二氧化硅薄片。关于这些蛋白质如何工作来创建复杂结构的一个假设是,残基在SDV内是保守的,不幸的是,由于Di的数量有限,很难识别或观察到这一点。
这些有机体之间的繁殖是通过二次分裂进行的,在此期间硅藻分裂成两部分,产生两个具有相同基因的新硅藻。每一个新的有机体都会收到双亲拥有的两个小果中的一个--一个大,另一个小,现在被称为上皮;并被用来构建第二个更小的小果,即假设囊。接受较大的锥体的硅藻变得与其亲本的大小相同,但接受较小的锥体的硅藻仍然比其亲本的小。这会导致硅藻种群的平均细胞大小减小。[17]然而,已经观察到某些分类群具有在不导致细胞大小减小的情况下进行分裂的能力。[37]然而,为了恢复硅藻种群的细胞大小,对于那些确实经历了尺寸减小的硅藻种群,必须进行有性繁殖和辅助孢子的形成。[17]。
硅藻的营养细胞是二倍体(2N),因此可以进行减数分裂,产生雄配子和雌配子,然后融合形成合子。受精卵脱落其硅质膜,长成一个被有机膜覆盖的大球体,即辅助孢子。在辅助孢子内形成一个最大尺寸的新的硅藻细胞,即初始细胞,从而开始新的一代。休眠孢子也可以作为对不利环境条件的反应而形成,当条件改善时就会萌发。[24]。
硅藻大多是不活动的;然而,在一些物种中发现的精子可以有鞭毛,尽管运动通常仅限于滑行运动。[24]在中心硅藻中,小的雄配子有一个鞭毛,而雌配子是大的,不活动的(卵配子)。相反,在羽状硅藻中,两个配子都没有鞭毛(同配子)。[17]某些蛛形目物种,即没有中缝的羽状硅藻(缝),已经被记录为异性配对,因此被认为代表中心和有中缝的羽状硅藻之间的过渡阶段。[37]。
海洋和湖泊中的某些种类的细菌可以利用水解酶分解有机藻类物质,从而加快硅藻在死亡和活着的硅藻中的溶解速度。[38][39][39]。
硅藻根据锥体的形状分为两类:中心硅藻和羽状硅藻。
羽状硅藻是两侧对称的。它们的每一个阀门都有开口,这些开口沿着凹槽呈狭缝状,它们的外壳通常平行于凹槽拉长。它们通过细胞质产生细胞运动,细胞质沿着纹路流动,总是沿着固体表面移动。
中心硅藻是径向对称的。它们由上瓣和下瓣-上皮和下膜-组成,每个瓣都由一个瓣膜和一条带状带组成,它们可以很容易地在彼此下面滑动,并随着硅藻的发展而膨胀,以增加细胞含量。中心硅藻的细胞质位于壳的内表面,并在位于细胞中心的大液泡周围提供中空的衬里。这个巨大的中央液泡由一种叫做细胞汁液的液体填充,这种液体类似于海水,但随特定离子含量的不同而不同。细胞质层有几个细胞器,如叶绿体和线粒体。在中心硅藻开始扩张之前,它的细胞核位于其中一个瓣片的中心,并在分裂完成之前开始向细胞质层的中心移动。中心硅藻有各种各样的形状和大小,这取决于贝壳从哪个轴延伸,以及是否有刺存在。
硅藻属于一个很大的类群,称为异形藻,既包括自养生物,如金藻和海藻,也包括异养生物,如水霉。异形石的分类仍然悬而未决:它们可能被指定为一个科、门、界或介于这些门、门、界之间的某种东西。因此,硅藻的等级从通常称为硅藻纲或硅藻门的纲到通常称为硅藻门的门(=门),其亚群的等级都有相应的变化。
据估计,现存的硅藻物种约有20,000种,其中约12,000种迄今已根据Guary,2012[41](其他来源给出的估计范围更广[17][42][43][44])被命名。已经描述了大约1,000-1,300个硅藻属,既有现存的,也有化石的,[45][46]其中约有250-300个硅藻属仅作为化石存在。[47]。
多年来,硅藻或硅藻纲(硅藻纲)或硅藻门(硅藻门)仅被划分为2目,分别对应于中心硅藻和羽状硅藻(中心硅藻和羽叶硅藻)。这种分类在1990年被round、Crawford和Mann作了广泛的修改,他们将硅藻划分为更高的等级(科,相当于动物学分类中的门),并将主要的分类单位提升为纲,保持中心硅藻为一个单一的纲,但将前一个羽状硅藻分为两个独立的纲,Fragilariophyceae和Silillariophyceae(后者保留了较旧的名称,但有一个修订的定义),它们之间包括45个目,其中大部分是Coscinodisophyceae,它们之间的分类包括两个独立的纲,即Fragilariophyceae和Silillariophyceae(后者保留了较旧的名称,但有一个修订的定义),它们之间包括45个目,大部分是。
今天(写于2020年年中),人们认识到,1990年的朗德等人的系统。随着较新的分子工作的出现,需要修改,然而,取代它的最佳系统尚不清楚,并且目前广泛使用的系统,如AlgaeBase、世界海洋物种登记及其贡献数据库Ditom Base,以及Ruggiero等人,2015年代表的所有生命的系统,都保留了round等人的系统。作为它们的基础,尽管将硅藻作为一个整体而不是作为一个纲/门来对待,为了更好地与系统学上相邻的群及其所含分类群的处理相一致,round等人的纲被降为亚纲。(有关参考,请参阅下面的各个部分)。
琳达·梅德林(Linda Medlin)和他的同事从2004年开始提出的一个建议是,将一些被认为与羽状硅藻关系更密切的中心硅藻目分离出来,作为一个新的纲,中生目,其本身与羽状硅藻的关系比其余的中心硅藻更紧密。这一假说-后来被命名为Coscinodiscophyceae-Mediophyceae-Bacillariophyceae,或Coscinodiscophyceae+(Mediophyceae+Bacillariophyceae)(Cmb)假说-已被D.G.Mann等人接受,他将其用作ADL中提出的硅藻分类的基础。等人的一系列合成(2005、2012、2019年),以及Archibald等人编辑的2017年原生动物手册中的硅藻章节,并进行了一些修改,反映了Medlin等人原始的Coscinodisophyceae的明显的非单系性。同时,E.C.Theriot领导的一个研究小组支持一种不同的系统发育假说,该假说被称为结构等级假说(SGH),并且不承认中体科为单系群,而另一项分析,即Parks等人,2018年的分析发现,径向中心硅藻(Medlin等人的Coscinodisophyceae)不是单系的,但支持中体科减去Attheya的单系。有关各方仍在继续讨论这些相互矛盾的方案的相对优点。[48][49][50][51]。
基于羽状硅藻在瓣膜上有或没有纵向凹槽的事实,称为中缝,[52]1990年按round,Crawford&Amp;Mann[42]进行的分类将硅藻(如硅藻)分为三类,中心硅藻(22目);无中缝的羽状硅藻(12目);和具中缝的羽状硅藻(11目),如下所示:[53]。
基于分子系统学,Medlin和Amp;Kaczmarska在2004年提出了硅藻的另一种分类,如下所示。[54]Medlin和同事为极地中心硅藻建立了一个新的类别,Mediophyceae(如果硅藻作为整体被列为一个纲而不是门,它可以被重新排序为一个亚纲),他们认为该硅藻与羽状硅藻而不是其他中心硅藻关系更密切,这一概念已经被一些人(例如Adl等人,2019年,见下文)所遵循和/或进一步修改,但不是所有随后的工作者在此时都遵循和/或进一步修改了这一概念。
在治疗的过程中。
.