树枝:莱昂纳多·达·芬奇的法则与生物力学模型(2014)
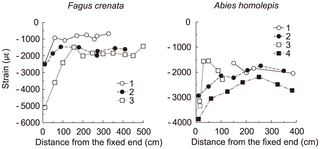
这项研究检验了莱昂纳多·达·芬奇的法则(即,在任何高度的分支点之上的所有树枝的横截面积的总和等于树干或分支点正下方的横截面积),使用基于两个生物力学模型的模拟:均匀应力模型和弹性相似模型。子代/母代比(即子代分支总横截面积与母代分支截面积在分支点的比例)的模型计算表明,当子分支的分支角和侧子分支的权重较小时,两种生物力学模型均符合达芬奇定律,但随着侧子分支的重量和(或)分支角的增加,模型偏离了达芬奇规则。这两个模型的计算值大致相似,但在某些方面有所不同。Fagus crenata和Abies homolepis的野外测量也符合这一趋势,其中随着侧枝相对重量的增加,模型偏离了达芬奇规则。然而,对于自然界中的分支模式,这种偏差很小,其中经验测量是在现实的测量条件下进行的;因此,达芬奇规则在真实分支模式的情况下并不与生物力学模型严重冲突,尽管模型计算描述了达芬奇规则和生物力学模型之间的矛盾。Fagus crenata的田间数据最符合均匀应力模型,表明应力均匀性是影响Fagus crenata枝条形态的关键因素,而不是弹性相似或达芬奇法则。另一方面,机械约束不一定对冷杉枝条的形态有显著影响,这取决于子枝的数量。相反,这些分支机构往往与达芬奇的规则是一致的。
许多研究检查了树的设计,这导致了几个经验规则。列奥纳多·达·芬奇提出,在任何高度的分支点之上的所有树枝的横截面积之和等于分支点正下方的树干或树枝的横截面积[1]。这种关系也可以用以下表述来表示:给定分支节点以下的分支横截面积等于该节点上方的子分支横截面积之和[2]-[6]。这一关系也可以表示为:给定分支节点以下的分支横截面积等于该节点上方的子分支的横截面积之和[2]-[6]。这就是众所周知的莱昂纳多·达·芬奇法则,或面积保持法则[2]。然而,并不是所有的分支机构都符合达芬奇的规则。Sone et al.。[4]发现一个枝条横截面积的年均生长量小于其子枝生长量的总和。这是因为当年生长面积占树枝横截面积的比例,在小而年轻的树枝中几乎总是比在大而老的树枝中更大。作者指出,如果基瓣增长的下降每年都在重复,那么达芬奇的规则就不会成立。Sone et al.。[6]证明了在宏碁,只有经历过脱落的树枝才遵循达芬奇的规则。
其中一个原因是不同意达芬奇的规则在一些分支可能是表达在他们的生物力学结构方面。对于树木来说,在形态形成过程中抵御诸如重力之类的机械应力是生存所必需的[7]-[9]。机械应力的影响大小随树形的不同而不同。因此,流行的观点一直认为树枝会自我调整以适应其机械约束,这导致了试图分析树木的生物力学设计。的确,机械环境的变化确实会影响树木的形态。用绳子系在适当位置以减少风力的机械刺激的树木比对照样本长得更高[10]。此外,当树木的机械安全性较低时,安全系数,即临界屈曲高度(由树干直径估计)与实际树高之间的比率较小;生长在密林中受保护条件下的树木的安全系数低于生长在开阔环境中的树木,后者暴露在更强的风力下[11]。这些关系表明:(I)存在改变树干或树枝直径和长度之间关系的生物力学限制(反映了机械安全性),以及(Ii)树枝生物力学安全性的大小应给予适当的值。树木的水力阻力也会影响其高度生长[12]。Taneda和Tateno[13]比较了机械和水力限制,得出结论,被子植物和裸子植物当前枝条中生物量的分配主要受机械支持标准的制约(尽管在某些情况下
在我们的模拟中,我们考虑了一个水平分支在一个水平面内分成n个子分支,并对n为2或3的虚拟分支进行了计算,假设分支主要受其自身重量的影响。每个女儿都由I(I = A,B等;图1)标识。假定分支的应用载荷仅由其自身重量组成,其作用方向垂直于分支所在的平面。我们在假设分支具有恒定的安全系数(均匀应力)的基础上,计算了分支点处的子分支和母分支的直径。然后我们计算了女儿/母亲的比例。
在该模型中,将分支视为在竖直方向上受力的水平梁。根据材料力学,作用在任意点的梁的直径与载荷之间的关系可表示为(1)[22],其中M是由载荷的大小和距加载点的距离确定的弯矩,σx max是最大弯曲应力。在均匀应力模型条件下,σx max为常数,直径仅与力矩有关。
发生在母子分支(分别为M,M和MI)分支点处的力矩为(2)(3),其中Wi是子体i的重量,gi是子体i的重心到分支点的距离,θi是子体i的分支角。在计算子体/母体比时,可以将直径测量点设置为分支点。但是,在野外测量时,分支点和测量点之间可能会有一些距离。考虑到这一点,力矩可以改写为(2‘)(3’),其中m M和m i分别是分支点和母子i的测量点之间的距离。在公式(2‘)中,假设分支点和母枝上测量点之间的截面对M影响不大。我们对m M和mi采用零和一些概率值(1、2、3和5 cm)进行计算。将公式(2’)或(3‘)代入(1)可得到母体和子体的直径。应用这些直径,可以从(4)中得到母体在分支点的横截面积,其中dM是母体分支的直径。子体的总横截面积可由(5)求得,其中di为子体i的直径。根据公式(4)和(5),根据方程式(6),子体/母体比为(6),子体/母体比与最大弯曲应力无关。
我们计算了不同分枝角度和不同子代重量情况下的子母比。角度0.1°~90°,子代体重1~10 kg。在n等于3的分支的模拟中(见下面的结果),主子分支(与图1中所示的分支B一样)的权重被设置为10千克。
在弹性相似假设中,假定树枝顶端的挠度与树枝的总长度成正比[8]。换句话说,分支的任意微截面(具有恒定的相对长度)的偏转角度由其在分支上的相对位置确定,与分支长度无关。
McMahon和Kronauer[8]假设锥形梁在某一点的垂直平面直径与s3/2成正比,其中s是从(选定的)点到顶端的分支截面的长度。King和Loucks[14]也引用了弹性相似模型,并解释了在自身重量下分支弯曲的直径和长度之间的关系:对于保持自相似的分支,分支重量W与d2l成正比,其中l是分支的长度,因为d2l与分支体积成正比。力矩臂与分支长度成正比。因此,(7)如果分支保持弹性相似的形式,则曲率与长度之间的关系可记为(8),其中r是分支在其底部的曲率半径。根据悬臂梁理论[22],(9)将(9)和(7)代入(8),直径与长度的关系为(10)。在本研究中,我们使用了可由上述表达式得到的异速生长关系(A)(B)(C),其中k1,k2,k3为常数,g为重心至分支底座之间的距离。
现在我们考虑一个没有分叉的水平直枝,它的形状类似于McMahon和Kronauer[8]所描述的形式。如果支管锥度的确定遵循弹性相似原则,则应严格保持支管任意截面的偏转角。根据CAN
将公式(12)和(15)代入(13),将(s l+m M)设为s(: = ηλ),得到保持常数dθ/dη:(16)所需分支点处的母枝的d4,其中s l是最长的子枝的长度。方程(16)的平方根为dM2,可用于计算AMM( = πdM2/4)。
子枝的横截面面积之和是(17)式(17)、(16)和(A),将(si−mi)设为s,子枝/母比表示为(18),其中si是每个子枝的长度。代入方程式(2‘)、(B)和(C),方程式(18)可排列为(19)。利用方程式(19),我们计算了具有各种分支形式的虚拟水平分支的子母比。对于k3,我们使用异速生长回归方程得到的值表示树枝长度与树枝重心到分枝点的距离之间的关系,这两种冷杉的树枝长度与分枝点之间的距离与树枝的重心距离之间存在着一定的关系。这些经验数据是通过下一节描述的方法获得的。如果任何分支形式的k3为常数,则子代/母体比随侧向子代的长度和分枝角度的不同而不同。
从生长在日光植物园(日光,Tochigi,Japan,E139°37‘,N36°45’)的日光植物园(Nikko,Tochigi,Japan,E139°37‘,N36°45’)的日光植物园(Nikko,Tochigi,Japan,E139°37‘,N36°45’)的日光植物园(Nikko,Tochigi,Japan,e139°37‘,N36°45’)的下部树枝(约30年生)和冷杉(年龄不详)标本中收集了几个树枝的直径、这些树节的远端部分的长度、远端部分的重心与测量部分之间的距离以及每个截面上每个树枝重量所产生的力矩的数据。随后,我们用指数回归的方法研究了每个物种的远端部分到测量部分的重心距离和长度之间的关系。我们用最小二乘法对曲线进行拟合。在上述模型计算中使用了回归曲线方程。
我们还收集了母子树枝的直径、女儿的分枝角度,以及使用同一棵树上其他较低的树枝从母树枝上分枝的女儿数量的数据。测定了6株江淮水青树和7株冷杉的13根树枝。取样树高分别为14.1~16.9m(14.7±1.3m,平均±SD)和9.4~22.6m(13.8±5.5m)。胸高平均直径分别为(26.4±1.5)cm和(31.6±13.1)cm。分支附加到。
.